![]()
![]()
![]()
Глава IV.12.
Обмен липидов
Переваривание и всасывание
Триацилглицериды (ТАГ)
или просто жиры поступают в организм с пищей животного и растительного
происхождения. В большом количестве они содержатся в сале, растительном и
сливочном масле, мясе, куриных яйцах, печени.
В ротовой полости и желудке эти процессы не идут из-за отсутствия ферментов.
В 12-перстную кишку тонкого
отдела кишечника с соком поджелудочной железы поступает липаза
в виде неактивной формы – пролипазы. С желчью туда же поступают желчные
кислоты, под действием которых липаза активируется.
Желчные кислоты
ориентируются на каплях жира, что приводит к уменьшению поверхностного
натяжения и дроблению их на более мелкие. Т.е. желчь эмульгирует жиры. На
поверхности таких мелких капель адсорбируется липаза и гидролизует эфирные
связи в молекулах триацилглицеридов. В результате от глицерина отщепляются
поочередно остатки жирных кислот (ЖК).
Высвобождающиеся ЖК усиливают эмульгирование жиров. Желчные кислоты
образуют комплекс с ЖК и моноацилглицеридами,
который легко проникают в клетки слизистой оболочки кишечника. В толще
слизистой желчные кислоты отщепляются от ЖК и с портальным кровотоком поступают
обратно в печень, где вновь включаются в состав желчи.
Из кишечника ЖК транспортируются по лимфе и крови к органам и тканям.
Поскольку эти вещества гидрофобны, то они переносятся по крови в комплексе с
белками, образуя липопротеиды. Наиболее распространены: хиломикрон и
липопротеиды очень низкой плотности (ЛПОНП).
Окисление
жирных кислот
ЖК во многих
тканях, в первую очередь в скелетных мышцах и миокарде вовлекаются в процесс b-окисления. Он протекает в митохондриях и
начинается с активации, т.е. присоединения КоА к ЖК с
образованием
RCOOH +HSKoA+АТФ
= RCO-SКoA+АМФ.
Мембрана митохондрий
непроницаема для ЖК даже в активированной форме, поэтому ацил-КоА соединяется
со специальным переносчиком карнитином.
Образуется ацилкарнитин,
который проникает в митохондрии, где вновь распадается на карнитин и ацил-КоА.
Процесс b-окисления включает 4 стадии
(Рис. 4.12.1.).
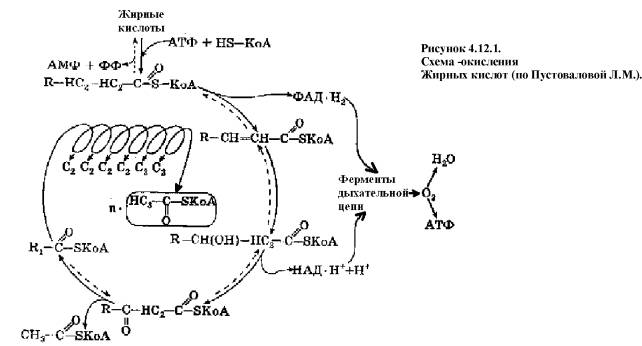
- дегидрирование ацил-КоА до дегидроацил-КоА
с участием ФАД-зависимой дегидрогеназы;
- присоединение к дегидроацил-КоА воды в b-положении с
образованием гидрооксиацил-КоА
при участии гидратазы;
- дегидрирование гидрооксиацил-КоА до b-кетоацил-КоА
при участии НАД-зависимой дегидрогеназы;
- расщепление тиосвязи при
участии тиолазы с образованием ацил-КоА и ацетил-КоА.
В реакциях дегидрирования на
1 и 3 стадиях образуются восстановленные коферменты, которые передают атомы
водорода на ЭТЦ, где синтезируется АТФ.
Каждая ЖК при очередном
цикле распадается на ацил- и ацетил-КоА, при этом цепь укорачивается на два
углеродных атома. Ацетил-КоА поступает в ЦТК, а ацил-КоА вновь проходит все 4
стадии. Это повторяется до тех пор, пока при очередном цикле не останется
ацетил-КоА. Из ЖК образуется ацетилов в два раза меньше, чем количество атомов
углерода в ней.
Окисление
ненасыщенных ЖК происходит так же, как и окисление ненасыщенных ЖК. Существует
дополнительный фермент трансеноилизомераза, который осуществляет перемещение двойной
связи из положения 3-4 в положение 2-3. При этом получается нормальный
субстрат, который может подвергаться дальнейшему окислению, как и ЖК с четным
числом атомов.
Окисление ЖК с
нечетным числом углеродных атомов
Они редко встречаются в
природе, но образуются в ходе окислительного расщепления валина и изолейцина.
Окисление происходит по обычному пути, до тех пор, пока в конце очередного
цикла не останется трехуглеродный фрагмент в виде пропионил-КоА.
Он далее подвергается ферментативному карбоксилированию в результате которого
получается метилмалонил-КоА,
который затем изомеризуется в сукцинил-КоА.
Под воздействием сукцинилтиокиназы из него
образуется сукцинат и
ацетил-КоА.
Кетоновые тела
Печень обладает способностью
ферментативным путем превращать ацетил-КоА в свободную ацетоуксусную
кислоту и эфир b-оксимасляной
кислоты. Они доставляются кровью к периферическим органам и тканям,
где включаются в ЦТК. В норме кетоновых тел содержится немного. Увеличение
этого показателя наблюдается при голодании и сахарном диабете и называется
кетоз. В крови – кетонемия, в моче – кетонурия.
Биосинтез ЖК
Протекает в
цитоплазме и включает следующие реакции:
1.
карбоксилировние
ацетил-КоА до малонил-КоА (Е: карбоксилаза, кофермент - биотин);
2.
соединение
ацетил-КоА и малонил-КоА с ацетилпереносящими белками;
3.
конденсация
ацетил-КоА и малонил-КоА с образованием комплекса ацетоацетил-ацетилпереносящий
белок;
4.
восстановление
кетоновых групп до спиртовых (кофермент - НАДН);
5.
отщепление
воды с образованием ненасыщенной связи;
6.
насыщение
двойной связи, при этом образуется бутирил-КоА (кофермент - НАДФН).
Бутирил-КоА вступает в новый
цикл, где удлиняется на 2 атома. Циклы повторяются до получения необходимой
длины цепи.
Синтез сложных липидов
Триглицериды синтезируются
на основе глицерофосфата
и ацил-КоА. Вначале присоединяется два
ацила, а затем под действием фосфатазы теряется
остаток фосфата и присоединяется третий ацил.
Биосинтез стероидов
Холестерол синтезируется на
основе ацетил-КоА. Около 80% в печени, 10 % в клетках кишечника и 5% в клетках
кожи. Желчные кислоты образуются в печени из холестерола.
ЛИТЕРАТУРА К ГЛАВЕ IV.12.
1. Бышевский А. Ш., Терсенов
О. А. Биохимия для врача // Екатеринбург: Уральский рабочий, 1994, 384 с.;
2. Кнорре Д. Г., Мызина С.
Д. Биологическая химия. – М.: Высш. шк. 1998, 479 с.;
3. Ленинджер А. Биохимия. Молекулярные основы
структуры и функций клетки // М.: Мир, 1974, 956 с.;
4. Пустовалова Л.М. Практикум по биохимии //
Ростов-на Дону: Феникс, 1999, 540 с.
5. Методы биохимических
исследований (липидный и энергетический обмен). Под ред. М.И.Прохоровой // Л.:
Изд-во Ленинградского университета, 1982. - 327 с.;
6. Ньюсхолм Э., Старт К.
Регуляция метаболизма. Под ред. Э.Г.Ларского. - М.: Мир, 1977. - 407 с.
![]()
![]()
![]()