![]()
![]()
![]()
Глава IV.14.
Взаимосвязь обменов
Обмен веществ складывается из химических превращений различных по природе веществ. Но можно назвать несколько общих целей метаболизма: генерация энергии и восстановленных эквивалентов, образование строительных блоков для биосинтеза.
Генерация энергии
Из различных по химической природе веществ
высвобождается энергия их химических связей в результате ряда
окислительно-восстановительных реакций. Катализируют такие реакции ферменты
класса оксидоредуктаз, который насчитывает 17 подклассов. Восстановленные коферменты
транспортируют электроны к единой структуре, обеспечивающей постепенное
высвобождение и запасание энергии, – электронтранспортной цепи. Конечным
акцептором этой цепи является молекулярный кислород, который доставляется
посредством оксигемоглобина. Конечными продуктами длительного и
многоступенчатого процесса генерации энергии являются макроэргические
соединения (АТФ, УТФ и др.), вода и энергия в виде тепла.
Генерация восстановленных эквивалентов
В результате окислительно-восстановительных реакций,
происходящих на этапах углеводного или липидного обменов, образуются
восстановленные коферменты (НАДН, НАДФН, ФАДН, ФМНН), которые затем могут
использоваться как общие для всех метаболических путей доноры водорода.
Образование строительных блоков для биосинтеза
Строительные блоки возникают как промежуточные
продукты процессов катаболизма, ведущих к образованию энергии и восстановленных
эквивалентов. Следовательно, все эти этапы объединены в единый
многофункциональный процесс, направленный на поддержание жизнедеятельности
клетки и постоянное обновление ее структур.
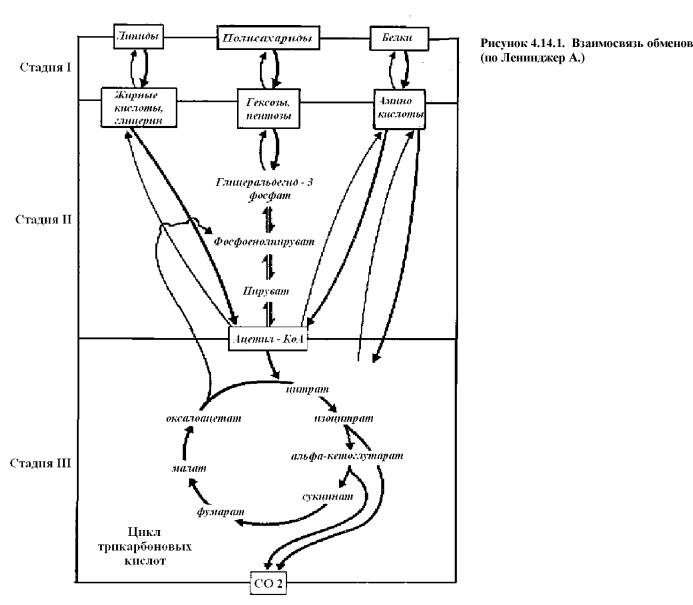
Объединение нескольких метаболических путей в единый
процесс неизбежно приводит к возникновению общих промежуточных метаболитов
(рис. 4.14.1.).
Глюкозо-6-фосфат образуется из глюкозы и
гликогена. Может расходоваться на:
- синтез глюкозы и гликогена;
- гликолиз до образования пирувата, у которого также несколько путей
использования;
- поступать на ПФП и превращаться в рибозо-5-фосфат.
Пируват образуется в результате
гликолиза, или в ходе превращения липидов, а также из аланина (реакцией
переаминирования). Превращается
- в лактат;
- в аланин (синтез белка);
- в оксалоацетат (глюконеогенез);
- в ацетил КоА (ЦТК).
Ацетил-КоА образуется из пирувата (а,
следовательно, аланина, глицерола) и ЖК. Поступает:
- в ЦТК;
- используется при бетта-окислении ЖК.
Общие механизмы регуляции обменных процессов
Скорость метаболических превращений зависит в большинстве случаев от концентрации и активности лимитирующих ферменов. Они еще называются регуляторными и катализируют обычно необратимые реакции. Активность таких ферментов как правило регулируется концентрацией одного из последующих продуктов или конечного продукта данного метаболического пути. Так при накоплении АТФ угнетается активность фосфофруктокиназы – ключевого фермента гликолиза, т.е. клетке на данный момент достаточно макроэргов. Активируется этот фермент при накоплении АДФ и АМФ.
Активность ферментов в свою очередь напрямую зависит
от концентрации коферментов (витаминов) и кофакторов (минеральных веществ). Так
дефицит витамина В1 (ТДФ) тормозит реакции окислительного
декарбоксилирования альфа-кетокислот. При этом замедляются отдельные этапы
гликолиза, ЦТК, тормозится окисление ЖК, химическое превращение АК и синтез
белка, возникает дефицит НАДФН, который образуется в пентозофосфатном шунте.
Снижение концентрации общих метаболитов сказывается
на уровне обменных процессов во всей клетке. В то же время, уровень таких
метаболитов может восполняться за счет различных биохимических реакций.
Т.о.
три вышеописанных элемента, составляющих общую цель метаболизма, объединены
общими механизмами регуляции:
1)
наличием
общих лимитирующих реакций и ферментов;
2)
аллостерическим
характером регуляции ключевых ферментов;
3)
принципиально
одинаковым механизмом гормональной регуляции;
4)
наличием
общих метаболитов;
5)
возможностью
быстрого переключения с одного метаболического пути на другой, за счет наличия
общих метаболитов.
Гормональная регуляция обмена веществ
Тесная взаимосвязь обменов, а именно углеводного, липидного и белкового на уровне всего организма обеспечивается четкой регуляцией со стороны эндокринной системы. Поэтому многие гормоны стоят на пересечении этих трех обменов.
Инсулин – снижает уровень сахара в
крови.
Углеводный обмен:
1) активирует транспорт глюкозы через клеточную
мембрану;
2) ускоряет окислительный распад глюкозы (активирует
ферменты ЦТК);
3) ускоряет гликогеногенез в печени и мышцах;
4) тормозит гликогенолиз и глюконеогенез.
Липидный обмен:
1)
ускоряет
липогенез из продуктов распада сахаров;
2)
замедляет
липолиз.
Белковый обмен:
1)
ускоряет
протеиногенез из продуктов распада сахаров.
Панкреатический глюкагон – увеличивает содержание сахара в крови.
Углеводный обмен:
1)
ускоряет
распад гликогена и глюконеогенез в печени;
2)
ограничивает
активность гликогенсинтетазы под действием инсулина (прямой антагонист).
Белковый обмен:
1)
тормозит
синтез белка;
2)
ускоряет
протеолиз;
3)
уменьшает использование глюкозы в процессе синтеза АК.
Адреналин – повышает уровень сахара в
крови.
Активирует гликогенолиз.
ЛИТЕРАТУРА К ГЛАВЕ IV.14.
1. Бышевский А. Ш., Терсенов О. А. Биохимия для врача // Екатеринбург: Уральский рабочий, 1994, 384 с.;
2. Ленинджер А. Биохимия. Молекулярные основы
структуры и функций клетки // М.: Мир, 1974, 956 с.;
3. Методы биохимических
исследований (липидный и энергетический обмен). Под ред. М.И.Прохоровой // Л.:
Изд-во Ленинградского университета, 1982. - 327 с.;
4. Ньюсхолм Э., Старт К.
Регуляция метаболизма. Под ред. Э.Г.Ларского. - М.: Мир, 1977. - 407 с.
![]()
![]()
![]()