![]()
![]()
![]()
Глава IV.7.
Биологические мембраны
Клеточная мембрана - это неотъемлемый компонент
любой клетки. Ее роль в первую очередь состоит в том, чтобы отграничить
внутреннее пространство клетки от внешней среды, а у эукариот, кроме того,
разделить внутреннюю часть клетки на функционально значимые отсеки: ядро и
митохонодрии. Во внешней клеточной мембране (цитолеме) функционируют
транспортные белки, рецепторы. Структурной основой всех мембран являются
липиды, в частности фосфолипиды, представленные двумя соединениями
фосфодиацилглициринами и сфингомиелинами.
Фосфотидилглицерин - наиболее распространенный
представитель фосфолипидов он присутствует в мембранах всех живых организмов и
является производным фосфатидной
кислоты. В основном это кислоты с длиной углеродной цепи 12-24
атома, либо полностью насыщенные либо имеющие одну или несколько несопряженных
двойных связей. В структуре фосфолипидов заложена важная функциональная
особенность: содержание длинной гидрофобной цепи и гидрофильной
"головки" из фосфатидной кислоты.
Сфингомиелины являются производными аминоспирта сфингозина.
Они характерны только для мембран животных клеток.
Наряду с этими компонентами мембраны содержат белки
и связанные с ними углеводы.
Мембраны представляют собой плоские образования
толщиной в несколько молекул (60-100 Å). Основу составляет липидный
бислой, где гидрофильные головки обращены к воде внутри и снаружи клетки, а
гидрофобные хвосты из жирных кислот как бы выталкиваются из воды и сливаются
между собой внутри мембраны (рис.
4.7.1.). Отдельные участки мембраны, липиды которых содержат больше насыщенных
ЖК находятся в жестком состоянии, другие, где содержится больше ненасыщенных
ЖК, в более расплавленном. Между
ацилными цепями липидного бислоя содержится холестерол, он
препятствует их кристаллизации, т.е. поддерживает состояние текучести.
Мембрана не статическое образование, а благодаря
жидкокристаллической структуре она является двухслойным раствором, в котором
липиды способны диффундировать как параллельно поверхности мембраны, так и из
одного монослоя в другой.
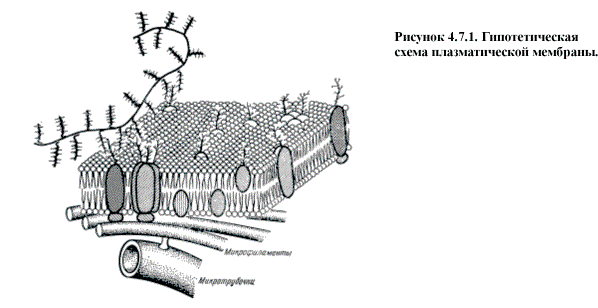
В структуру мембран обязательно входят белки и их
состав варьирует в зависимости от функции той или иной мембраны. В зависимости
от прочности связи с мембраной различают периферические и интегральные белки. Интегральные
белки располагаются между липидами монослоя или пронизывают весь
бислой, часто возвышаясь над поверхностью мембраны. Периферические белки
связаны с мембранами электростатическими и водородными связями и часто
взаимодействуют таким образом с интегральными белками (рис. 4.7.1.).
Белки выполняют следующие функции:
1)
транспорт
(трансмембранный перенос веществ);
2)
преобразование
энергии (ферменты дыхательной цепи);
3)
коммуникативную
(рецепторные белки связывают клетку с окружающей средой);
4)
и
ряд специфических функций.
Трансмембранный
транспорт веществ
Липидный бислой практически непроницаем для ионов и большинства
полярных молекул. Исключение составляет вода. Перенос веществ через клеточную
мембрану осуществляется одним из трех путей: простой диффузией, облегченным и
активным транспортом.
Простая диффузия осуществляется за счет
теплового движения молекул из зоны с большей концентрацией в зону с меньшей
концентрацией. Разность концентраций по обе стороны мембраны называется
градиентом концентраций. При переносе заряженных частиц важную роль играет
разница потенциалов. Обычно внутри клетки накапливаются отрицательно заряженные
частицы. Таким путем транспортируются: Н2О, СО2, О2.
Перенос веществ простой диффузией прекращается когда концентрация вещества с
одной и с другой стороны мембраны выравнивается.
Облегченная диффузия более распространенный
способ транспорта веществ через мембрану. Здесь принимают участие специальные
белки-переносчики. К ним относятся ферменты транслоказы
и пермиазы. Они связывают своим
активным центром вещество с одной стороны мембраны и переносят его сквозь
гидрофобный слой мембраны на ее другую поверхность. Еще один вариант такой
диффузии: после присоединения транспортируемого вещества меняется конформация
белка-переносчика и в мембране открывается специальный гидрофильный канал, по
которому и проникает вещество.
Активный транспорт - это транспорт против
градиента концентраций и происходит при затрате энергии. Если источник энергии
АТФ, то это первично-активный транспорт, если энергия получается за счет
переноса в этот момент другого вещества по градиенту концентраций - это вторично-активный
транспорт.
Типичный первично-активный транспорт это калий-нартиевый насос. Он локализован в
плазматической мембране практически всех клеток и переносит ионы калия и натрия
против градиента концентраций с использованием энергии АТФ (рис. 4.7.2.).
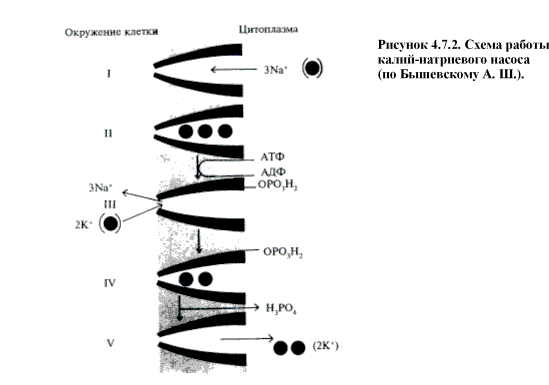
Собственно насос это фермент Na+-K+-АТФ-аза, интегральный белок
пронизывающий липидный бислой мембраны насквозь. Внутри клетки к активному
центру этого фермента присоединяется 3 иона Nа+, при этом
фермент активируется и расщепляет АТФ на АДФ и остаток фосфорной кислоты. Этот
остаток присоединяется к самому ферменту и изменяет его пространственную
конформацию. При этом с внутренней стороны мембраны закрывается ионный канал,
но открывается на наружной поверхности. Ионы Na+ отсоединяются от фермента,
но в это же время к другому активному центру
фермента присоединяются 2 иона К+. Это вновь изменяет его
пространственную конформацию, отщепляется остаток фосфорной кислоты и
открывается канал для проникновения К+ внутрь клетки. Далее цикл
повторяется. Т.о. при выносе из клетки 3 ионов Na+ в нее проникает 2 иона К+.
Это ведет к появлению электрического потенциала, который называется
трансмембранным электрохимическим потенциалом. Если насос прекратит работу, то
ионы Na+ и К+ начнут перемещаться в обратном
направлении. По такому же механизму транспротируются ионы Са2+
(фермент Са2+-АТФ-аза).
Перекисное окисление липидов мембран
Перекисное окисление липидов является
свободнорадикальным процессом, инициация которого происходит под действием
активных форм кислорода. К ним относятся супероксид-ион О2-,
пероксид-ион О22-
(из Н2О2), гидроксильный радикал НО*, гидропероксидный
радикал НОO* .Они образуются в результате процессов окисления
протекающих в клетке при участии железо-содержащих ферментов (цитохромов).
Свободные радикалы это частицы с неспаренными
электронами на внешних орбиталях

Механизм инициации ПОЛ
1) R - CH - … + OH* = R - C* - или (R*)
свободный радикал высшей жирной кислоты (ВЖК);
2) R* + O2 = R - O - O*
пероксидный ион ВЖК;
3) R - O - O* + R-СH = R - O - OH (гидроперкеись ВЖК) + R*.
Реакции 2 и 3 теперь могут
идти без активных форм кислорода они превращаются в цепные.
Продукты перекисного окисления
липидов:
R* свободный радикал ВЖК,
R-О-О* пероксидион ВЖК,
R-О-ОH гидроперекись ВЖК,
НС=О - СН2 - С=ОН малоновый
диальдегид,
R-CH = CH - CН2 - СН = СН2
диеновые
коннъюгаты.
Активация процессов ПОЛ, независимо от факторов
индукции, может вести к деструктивным изменениям в клетках, что связано с
накоплением продуктов, способных инактивировать мембранные ферменты, нарушать
белок-липидные взаимодействия в мембранах, образовывать межмолекулярные
сшивки, изменять вязкость липидной
фракции, что препятствует образованию фермент-субстратного комплекса.
Но, несмотря на вышеперечисленные патологические
процессы, важно отметить, что ПОЛ это физиологический процесс и он имеет важное
значение для организма. Уровень продукции ПОЛ в норме контролируется рядом
антиоксидантных веществ и ферментов. К ним относятся витамины: Е,
С,
бета-каротин, убихинон (коэнзим Q) и антиоксидантные
гем-содержащие ферменты супероксиддисмутаза (СОД), каталаза, глутатионпероксидаза,
глутатионредуктаза. Высокая активность ПОЛ может привести к подавлению
активности антиоксидантных ферментов. В этом случае в клетках развиваются
вышеописанные процессы, которые с клеточных мембран переходят на
цитоплазматические структуры. Это приводит к денатурации клеточных белков,
снижению активности ряда ключевых метаболических ферментов и повреждению
клеточного генома. Такое явления носит название окислительный стресс. Все
это заканчивается гибелью клетки по пути некроза (разрушения клеточных
структур) или апоптоза (запрограммированного суицида).
ЛИТЕРАТУРА К ГЛАВЕ IV.7.
1. Бышевский А. Ш., Терсенов
О. А. Биохимия для врача // Екатеринбург: Уральский рабочий, 1994, 384 с.;
2. Кнорре Д. Г., Мызина С.
Д. Биологическая химия. – М.: Высш. шк. 1998, 479 с.;
3. Ленинджер А. Биохимия. Молекулярные основы структуры
и функций клетки // М.: Мир, 1974, 956 с.;
4. Пустовалова Л.М. Практикум по биохимии //
Ростов-на Дону: Феникс, 1999, 540 с;
5. Gutteridge J.V.C., Halliwell B. The
measurement and mechanism of lipid peroxidation in biological systems //
Trends in Biochem. Sci., 1990, р. 129-135.
![]()
![]()
![]()